16 okt. 2018
8 minuten
Planten kunnen niet weglopen als hun omgeving op een negatieve wijze verandert. Verrode golflengten rond de 700 nm zijn essentieel om te overleven. Voor kwekers biedt dit licht tal van praktische toepassingen door in te spelen op het fytochroomsysteem. Een lesje manipulatie door groene vingers met rood licht.
Door Johann Waldherr en Richard Blakey, Würth Elektronik
We weten dat de lichtvereisten van planten veel complexer zijn dan we oorspronkelijk dachten. Dit heeft geleid tot de ontwikkeling van verschillende ledtechnieken die een verscheidenheid aan lichtspectra produceren, zowel monochromatisch als polychromatisch. Sommige daarvan zitten nog in een experimentele fase. Een interessant gebied is het spectrum rond verrood licht. Dit betreft golflengten van 700 tot 800 nm, een gebied dat voor ons op de grens ligt van zichtbaar licht. Uit onderzoek blijkt echter dat deze golflengten resulteren in snellere groei, verhoogde biomassa en betere sensorische eigenschappen (zoals reuk, smaak, textuur en kleur). In tegenstelling tot mensen en dieren, kunnen planten niet bewegen. Het sedentaire bestaan van planten betekent dat ze zonder invloed van buitenaf, hun hele leven groeien en leven op dezelfde plaats. Dit lijkt een eenvoudige observatie maar het gevolg voor planten is dat ze moeten kunnen overleven als de omstandigheden van hun directe omgeving op nadelige wijze veranderen. Het omgaan met beperkte hulpbronnen zoals water, voedingsstoffen en licht, evenals dag-en-nachtcycli en jaarlijkse cycli, zijn van essentieel belang voor het overleven van de plant. Onderzoekers kunnen de manier waarop planten hiermee omgaan manipuleren zodat betere groeieigenschappen ontstaan.
Fotoreceptoren en het fytochroomsysteem
Licht is voornamelijk nodig voor fotosynthese, het voornaamste energie-omzettingsmechanisme van een plant, dat vooral te danken is aan rood en blauw licht via chlorofyl. Drie factoren zijn hier van belang:
- Lichtintensiteit: de hoeveelheid fotonen die de plant kan gebruiken;
- Fotoperiodiciteit: de blootstellingsduur;
- Lichtkwaliteit: de golflengten van licht waaraan de plant wordt blootgesteld.
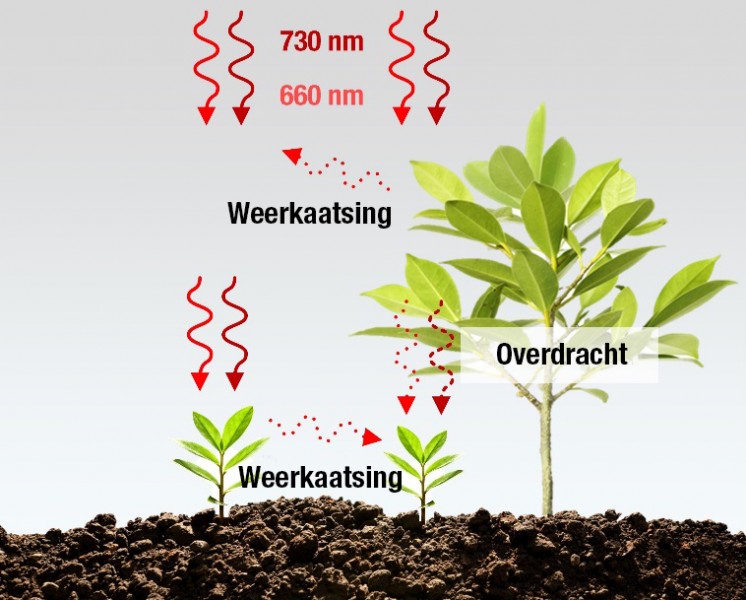
Maar licht heeft ook invloed op een aantal andere plantprocessen. Elk proces kan gekoppeld zijn aan een fotoreceptor die reageert op een specifiek bereik golflengten. Cryptochromen nemen blauw/UVA-licht waar en zijn verantwoordelijk voor fototropisme – naar het licht keren – en fotomorfogenese – het vormen van de plant onder invloed van licht – , terwijl fotoreceptoren genaamd fytochromen juist verrood licht waarnemen. Het fytochroomsysteem is in wezen afhankelijk van de interactie tussen twee golflengten en bestaat daarom uit twee soorten fytochromen die elk verschillende absorptiegolflengten hebben. Pr (fytochroom rood) heeft een absorptie van maximaal 660 nm en Pfr (fytochroom verrood) heeft een maximale absorptie van 730 nm. Interessant feit is echter dat Pr en Pfr hun moleculaire structuur omgekeerd kunnen inzetten, afhankelijk van de verhouding tussen de rode en verrode golflengten.
Pfr-fotoreceptoren worden beschouwd als de actieve vorm waarin de Pr-vorm verandert in de aanwezigheid van 660 nm rood licht. Pfr is fysiologisch actief, het initieert biologische reacties maar is onstabiel. Dit betekent dat in gevallen van verminderd of ontbreken van 660 nm licht, Pfr verandert in de Pr-vorm. De Pfr-vorm wijzigt ook in de inactieve Pr-vorm bij de aanwezigheid van 730 nm verrood licht. Daarom heeft het bereik (lichtkwaliteit) naast de blootstellingstijd (fotoperiodiciteit) en de totale hoeveelheid (lichtintensiteit) rode en verrode golflengten voor de plant een effect op het fytochroomsysteem. Verschillende verhoudingen van rode/verrode golflengten kunnen via biologische reacties een aanzienlijk effect hebben op de eigenschappen van planten.
Morfologische reacties op rood en verrood licht
Het fytochroomsysteem bestuurt verschillende moleculaire processen waardoor planten zich kunnen aanpassen aan de lichtomgeving. Deze reacties op rood en verrood licht hebben zich ontwikkeld als reactie op de omgeving en zijn in sommige gevallen overlevingsmethoden. Bijvoorbeeld om schaduw te vermijden voor gevallen waarin een plant niet genoeg licht krijgt. Dit kan ofwel het gevolg zijn van een vast object (dat al het licht tegenhoudt) of andere planten (die enkele lichtgolflengten doorlaten). Een andere reactie heeft betrekking op veranderingen in dagelijks en seizoenslicht, wat te maken heeft met de temperatuur- en luchtvochtigheidsschommelingen en de reactie van de plant op deze omstandigheden.
Lichtkwaliteit en het schaduwvermijdingssyndroom
In sommige ecosystemen kunnen planten in extreem hoge dichtheid groeien. In deze omgeving concurreren planten om een beperkte lichthoeveelheid, de bron voor fotosynthese. Deze concurrentie kan plaatsvinden tussen planten die op hetzelfde moment groeien of bij planten die groeien in de schaduw van een grotere plant. Om in deze omgeving te kunnen overleven, nemen planten de hoeveelheid schaduw waarin ze zich begeven waar via hun fytochroomsysteem. Daarbij zijn verschillende manieren waarop de hoeveelheid rood en verrood licht in een lichtconcurrerende omgeving de plant kan bereiken. De meerderheid van het licht in het verrode bereik reflecteert of komt via ander plantmateriaal bij de plant terecht. Door de weerkaatsing van omringende planten of doorgelaten licht onder een grote overkapping vermindert de verhouding rood/verrood licht dat de planten in de directe omgeving bereikt.
Dit betekent dat direct zonlicht een grote hoeveelheid rood licht bevat terwijl doorgelaten of weerkaatst licht van bladeren, geen rood licht bevat maar een relatief hogere hoeveelheid verrood licht. De rood-verrood ratio beïnvloedt het gedrag van het fytochroomsysteem en derhalve de schaduwvermijdingsreactie van planten die geen schaduw verdragen of veel van zonlicht houden. Talrijke onderzoeken tonen aan dat deze reacties gepaard gaan met een versnelde verlenging van de hypocotyl, het internodium (beide onderdeel van de stengel) en bladstengel, verhoogde horizontale bladhoeken en verminderde vertakking om zo meer zonlicht te vangen en fotosynthese mogelijk te maken. Dit gedrag moet ervoor zorgen dat de plant overleeft. Daarnaast kan de schaduwvermijdingsreactie ervoor zorgen dat de plant vroeg in bloei staat dat de groei vermindert en de voortplantingsfase start in een poging om te overleven.
De praktische toepassingen hiervan zijn afhankelijk van de gewenste eigenschappen van de groeiende plant. Bij sla bijvoorbeeld remt verrood licht het ontkiemen, maar het is belangrijk te weten dat de ontkiemreactie afhankelijk is van de laatste lichtbehandeling. Aanvulling met verrood licht gedurende de groeifase resulteert in verhoogde groei van scheuten en wortels met hoger vers scheutgewicht en bladoppervlak. Eucalyptusstekken hebben meer wortelgroei bij een lage verhouding rood tot verrood licht. De opbrengst van groene bonen is hoger bij verrood licht en het geeft tevens grotere tomatenplanten. Interessant feit is dat verrood licht ook stamgroei van planten remt waarbij het onwenselijk is dat ze te groot worden. In dit geval zijn groeiremmers minder of zelfs niet meer nodig.
Fotoperiodiciteit en dag-en-nachtcycli
Planten zijn tevens gevoelig voor de overgang van golflengten van rood naar blauw licht zoals gebeurt tijdens zonsopgang en in tegengestelde richting bij zonsondergang. Daarnaast zijn ze ‘bewust’ van het tijdstip waarop dit dagelijks gebeurt, wat van invloed is op plantprocessen zoals het bloeien. Zoals bij de meeste levensvormen moeten ook planten het natuurlijke ritme aanpassen aan perioden van activiteit en van inactiviteit om zo het energieverbruik te beperken tot de momenten waarop dit het efficiëntst en voordeligst is. Zo is het normaal gesproken niet voordelig voor een plant om ‘s nachts of in de winter te bloeien, aangezien er dan maar weinig of geen bestuivers actief zijn. Tijdens de plantontwikkeling is er een vegetatieve fase (groeien) en een voortplantingsfase (bloeien).
Fotosynthese zorgt er tijdens de groeifase voor dat planten, bladeren en scheuten groeien terwijl er bijna geen groei plaatsvindt in de bloeifase. Door de ontwikkeling van bloemen, kunnen planten worden bestoven en fruit of zaden produceren. Het proces van bloemproductie is onomkeerbaar zodra dit in gang is gezet. Daarom is de timing van cruciaal belang. De gerichte beïnvloeding van bloemproductie speelt een essentiële rol, met name voor sierplanten en planten die zaad produceren. Het fytochroomsysteem is het belangrijkste systeem voor het identificeren van en het inspelen op deze veranderingen. De plant produceert fytochroom Pr tijdens duisternis en vermeerdert deze. Overdag produceert de plant Pfr door verrood licht dat langzaam terugkeert in de inactieve Pr-vorm (half leven = 2,5 uur) tijdens het ‘duisternisterugkeer’ proces. Overdag verandert de Pr-vorm in Pfr, dat de Pr/Pfr-verhouding weer corrigeert. Hierdoor is tijdens perioden van duisternis de Pr/Pfr-verhouding hoog en overdag laag. De gevolgen van de reactie van het fytochroomsysteem op deze veranderingen is afhankelijk van het soort plant.
Er zijn zogenoemde lange-dagplanten (LDP) en korte-dagplanten (KDP). KDP hebben een donkere fase nodig om te bloeien die niet verstoord mag worden door licht. Zonder deze donkere fase zal de plant niet bloeien. LDP hebben juist een lichtfase nodig om te kunnen bloeien.
De dieprode led straalt 660 nm licht uit in het verrode bereik. De verrode led 730 nm in het donkerrode spectrum. Een plant die meer rood licht krijgt, in verhouding tot blauw licht, met een lagere rood/verrood-verhouding, zal dit interpreteren als de start van een nieuwe dag en mogelijk gaan bloeien. Als de verhouding toeneemt, weet de plant dat de overgang naar overdag is bereikt. Bij een afname van de verhouding, door de zonsondergang, ziet de plant minder licht en start met metabolische en morfologische veranderingen. De duur van de nacht heeft overigens ook veel invloed op de morfologie van de plant.
De praktische toepassingen hiervan zijn talrijk en afhankelijk van het soort plant. Kwekers van sierplanten kunnen dankzij de gerichte manipulatie van het fytochroomsysteem bepalen wanneer de producten klaar zijn om te verkopen. Dit is zeer nuttig als je kijkt naar de jaarlijkse vraagcycli voor verse sierbloemen. Dagen zoals Valentijnsdag en Moederdag, wanneer de vraag het hoogst is, vallen vroeg in het jaar en dus buiten de gewoonlijke bloeiperioden van de meeste sierplanten. Maar de plant kan tevens geforceerd worden om meerdere keren per jaar te bloeien. Bij gebruik van kunstlicht zijn de groei- en bloeicycli niet gebonden aan seizoenen. Daarom is er de mogelijkheid om planten te laten bloeien in de winter, wanneer de lichtomstandigheden laag zijn. Op dezelfde manier kunnen ongunstige bloeiperioden tijdens de zomermaanden voorkomen worden.
Voorbeeld: De modelplant Arabidopsis thaliana is een lange-dagplant en bloeit eerder bij lange-dagomstandigheden dan bij korte-dagomstandigheden. De chrysant is een kwalitatieve, korte-dagplant. Uniform bloeien is mogelijk bij een fotoperiode van 13,5 uur of korter. Als de fotoperiode langer is, treedt er geen bloei op. Bij het kweken van chrysant-stekken gebruikt de kweker tevens lange-dagomstandigheden gedurende ongeveer 2-3 weken voordat hij of zij overgaat naar korte-dagomstandigheden, gezien de vereiste stengellengte. Interessant feit is dat wanneer de nachtperiode wordt onderbroken door dieprood licht, het bloeien van de chrysant verminderd kan worden. Dit is tevens het geval voor planten uit de Perilla-familie en tarwe. Daarom kan de duur van de nachtperiode belangrijker zijn dan de duur van de lichtperiode. Het bloeien van de Kalanchoë, een korte-dagplant, wordt voornamelijk gecontroleerd door rood licht. Dankzij deze informatie kunnen kwekers hun eigen geoptimaliseerde lichtrecept ontwikkelen voor het kweken van planten.
Dit artikel komt uit Elektronica#11/12 , met meerdere artikelen over fotonica en Leds.
Anderen lazen ook